Most large land animals on Eurus belong to the clade Endostea, characterised by the presence of an internal skeleton. It is this skeleton in addition to their circulatory and respiratory systems that have allowed them to reach their large sizes.
Evolution of Endosteans
The evidence that previous researchers on Eurus have gathered points towards Endosteans descending from a group of insect-like organisms called Holocelypheans. Going further back, Holocelypheans descended from shelled mollusc-like organisms. On the course of their evolution towards a more insect-like form, these “molluscs” developed numerous hydrostatic legs under their body, and eventually grew a jointed exoskeleton over their whole body for greater protection.
The group of Holocelypheans ancestral to Endosteans, the Arthropterans, had wings and was capable of powered flight, shown in the image below. These wings originally developed from the second and rear pair of limbs, which were used for swimming by their aquatic ancestors. These limbs developed numerous hairs on them to increase drag and function as fins. Although the exact details are unknown, not long after they moved to land, these fins developed into wings
 | ||
The ancestral Arthropteran. This is the
group the Endosteans arose from. Although they look like Earth flies, they’re
more like flying spiders in a lot of ways
|

Arthropterans grew larger over time, but there a number of limitations to their size. One is the need to constantly shed their exoskeleton as they grow, leaving them weak and their weight relatively unsupported as their new cuticle hardens. One group of Arthropterans solved this by developing a layer of skin over their exoskeleton, which deposits new layers of skeleton over old ones as it grows. This is in stark contrast to their ancestors who grew their new exoskeletons underneath the last. Their old skeletons can be reabsorbed rather than shed. With this layer of flesh over it, their exoskeleton became an endoskeleton. This outer flesh consisted of not just skin but also fatty tissue, allowing the animal to hold as much fat as necessary without their skeleton restricting it.
Other adaptations included the development of a lung. This evolved from the tracheal system. In some species, the trachea can be expanded and contracted to allow for a kind of breathing, and it was this that allowed a lung-like organ to appear as the respiratory system further adapted to support a larger body. They also developed a closed circulatory system, rather than the open circulatory system of their ancestors.
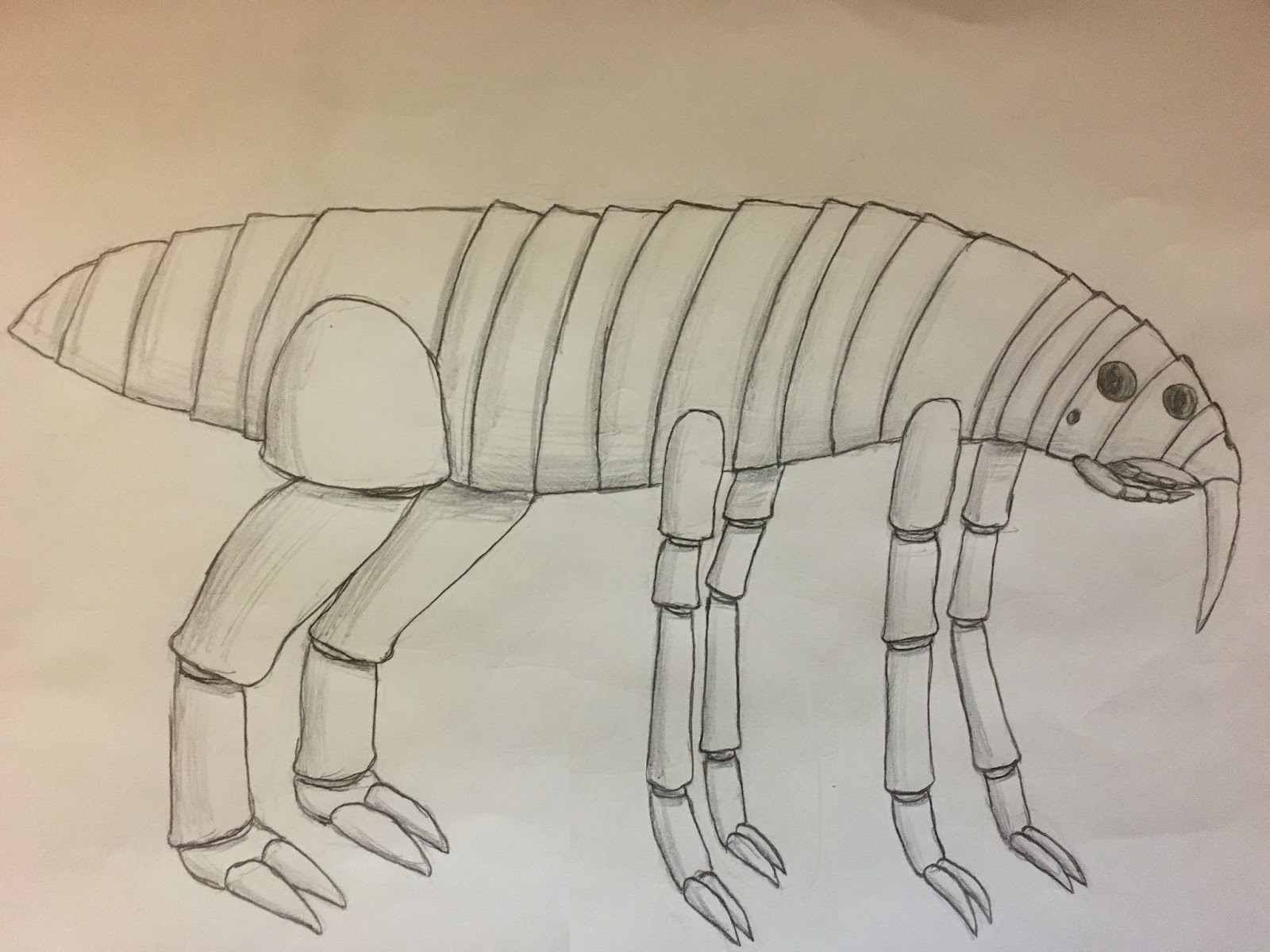
These adaptations allowed the first Endosteans to reach a much larger size then the insect-like organisms they descended from, a previously untapped niche on Eurus for land animals. The earliest Endosteans looked something like the animal below, although keep in mind this is just a reconstruction. Although the first Endosteans were octopedal, in most modern groups one or more of these pairs of legs was either lost or repurposed.
 |
| The ancestral Endostean |
Endosteans are split into two major groups; Platysomes and Tetrapterans. Platysomes tend to have flattened bodies to increase their ability to withstand strong winds, and they’ve lost all four wings. Their front legs have shrunk in size and came into use as jaws for feeding. In most lineages they are covered by a layer of skin that protects the mouth. Tetrapterans have retained their wings, and although many are incapable of flight they serve a function in temperature regulation and sometimes gliding. Tetrapterans have six legs, with the second to rear pair lost, although some groups have less limbs. Ancestrally they have two wing pairs, but in some groups one or both of these pairs have been lost.
In general, Platysomes have adapted to living out in open spaces in the wind, while Tetrapterans are more suited to enclosed environments like the planet’s “wind shadow forests”. However, both groups can be found in all environments.
Anatomy
 |
| Internal anatomy of an Endostean, showing the organs of the Platysome Monocerodon) |
 |
| Monocerodon skeleton |
Skeleton
In terms of structure, the skeletons of Endosteans are very similar to that of their ancestors, apart from the fact it’s internal.
At the torso and head, the skeleton consists of a series of overlapping plates, completely covering the internal organs but allowing for movement. In Tetrapterans, however, there are gaps between these plates, except at the head (where more protection is needed), sacrificing defence for lightness.
The bones of the limbs consist of hollow cylinders, with elastic connective tissue at each joint connecting these bones. The muscles are situated on the inside of these cylinders, rather than the outside, a remnant of their ancestry from animals with exoskeletons. They typically have six legs, at the end of each are a pair of claws. The bone of these claws is typically quite flexible, often consisting more of chitin and connective tissue than calcium carbonate, and they’re hollow, with muscle inside. This muscle is able to bend the claws, although in some groups the claws have hardened and become less flexible.
Tetrapterans have wings similar in structure to the legs. Each wing is supported by a single jointed appendage, with hollow cylindrical bones and muscles on the inside.
Nervous system
The central nervous system consists of a brain at the front of the animal, where most of the sensory organs are located, attached to a long spinal cord running along the underside of the body where it’s protected by bone.
Muscles
Muscles are located on the inside of the bones, and attach at their ends via connective tissue. Muscles consist of fibres aligned parallel to the long axis, as well as fibres arranged perpendicularly to the long axis, allowing them to both expand and contract, acting as a muscular hydrostat. This is probably a result of their ancestry from mollusc-like ancestors, whose entire bodies consisted of muscular hydrostats. This way, muscles control the animal’s movement by both pushing and pulling against bone. The insides of the bones have grove-like structures where the muscles attach to allow for this.
Circulatory and respiratory systems
Two hearts are present along the back. The frontmost heart is the lung heart, or anterior heart, a two chambered heart responsible for pumping blood through the lung. Further back is the systemic heart, or posterior heart, which can have up to four chambers and pumps blood throughout the body and back to the lung heart.
The systemic heart carries both oxygenated and deoxygenated blood, with oxygenated blood directly from the lung going though the upper atrium and ventricles, and deoxygenated blood going through the lower atrium and ventricles. The lung heart, however, only pumps deoxygenated blood, pumping it through its single atrium and ventricle.
The blood of endosteans contains the copper-based oxygen carrying protein haemocyanin, giving it its blue colour. Deoxygenated blood, however, is colourless. Endosteans don’t have the equivalent of red blood cells, with the haemocyanin instead suspended directly in their blood plasma rather than carried by cells.
Haemocyanin is only about a quarter as efficient as the haemoglobin of Earth vertebrates. However, it’s more efficient in the low oxygen and low temperature conditions found in the deep sea where the endosteans’ distant mollusc-like, and later crustacean-like ancestors lived. In fact, the molluscs and crustaceans of Earth use the same protein. This protein was retained throughout the evolution of holocelypheans on land and as endosteans evolved; however, the specific form of haemocyanin changed over time, and in modern endosteans a more efficient haemocyanin is used which is unknown on Earth. This haemocyanin is over twice as efficient as the usual kind in the conditions found on the surface.
Digestive system
In platysomes, the digestive system begins at a mouth near the front of the body, which is protected by a layer of flesh just under the head forming a pouch. The inside of this pouch can be accessed through an external sphincter, where food is taken in. Inside the mouth there are two extendible tongues, one above the throat and one below the throat, as well as two different jaw pairs used to masticate food. The jaws consist of the inner mandibles, which are smaller, not attached to the skull and rarely ossified, as well as the larger outer mandibles. The inner mandibles are used to grind food, acting like molars. The outer mandibles developed from the front limbs and are used to cut food, acting as incisors, both coming into contact with a chitinous “beak” at the front of the roof of the mouth that it cuts against.
The mouths of tetrapterans have some differences. Tetrapterans lack outer mandibles, with the inner mandibles possessing attachments to the skull and are used for cutting rather than grinding. The mouth is positioned more to the front of the head, rather than underneath, and it lacks the protective pouch of platysomes with the mandibles exposed. The upper and lower tongues are both present, although they tend to be retracted inside the throat. The anus also ends inside the throat rather than just below it, with only one hole visible from the outside. Unlike the inner mandibles of platysomes, tetrapteran inner mandibles are usually fully ossified.
Once food enters the throat it travels through the oesophagus, through a series of muscle contractions, where it eventually reaches a stomach. In most tetrapterans, there is a gizzard before the stomach; this allows food to be ground up before reaching the stomach, since the mandibles of tetrapterans don’t chew. These gizzards are lined with numerous hard chitinous plates and surrounded by a thick layer of muscle. The stomach breaks down food chemically with enzymes and acid, but plays very little role in absorption. The lower lobe of the stomach functions as a crop, storing food, but can be greatly reduced in some species. Both tubes connect to the upper stomach; food is able to pass entirely through the stomach without entering the crop. The stomach also plays a role in detoxification.
After being processed in the stomach, food enters the posterior intestines, which play some role in food absorption but mainly absorb water and filter blood. After having nitrogenous waste added to it the stool travels around to the anterior intestines where the majority of nutrient absorption takes place. As it approaches the front of the body it passes through the rectum, and then an anus, located near the throat. In platysomes, it is inside the mouth pouch just below the opening to the throat. The waste is then expelled out of the mouth.
Senses
 |
| Endostean eye anatomy |
Most major sensory organs are clustered near the front of the body. For vision, there are multiple paired eyes, usually between four and eight, as well as smaller ocelli in some groups. The major eyes are camera eyes able to resolve images, and although their general structure is comparable to that of the vertebrate eyes of Earth, on a closer look there are a number of differences.
Instead of a lens, the pupil consists of a small pin-hole that usually has a transparent protective membrane, with the light instead focused by a concave reflective structure called a speculum at the back of the eye. This focuses the image onto the retina, just behind the front of the eye, covered in photoreceptors that send signals down the optic nerve and to the brain. Images can be focused by changing the shape of the speculum, using muscles just beneath it pushing and contracting to modify its curvature as well as changing its position relative to the retina. The presence of the pupil in the middle of the retina creates a blind spot. There is also a second blind spot where the optic nerve passes through the retina.
The small ocelli developed much later, and are incapable of focusing or resolving a clear image. They likely appeared after the development of flight in holocelypheans, at least from the evidence I’ve been able to gather, and originated from photosensitive areas of the exoskeleton. They originally served a function in stabilising the flight of arthropterans, much like the ocelli of Earth insects do, but have come to assist the primary eyes in numerous other ways throughout endostean evolution. The ocelli are much harder than the primary eyes, consisting of a partially mineralised chitin lens with a retina at the back of the eye.
The hearing organs of endosteans are located near the front of the head inside the mouth. Each individual has a single “ear”, consisting of a loose bone able to respond to vibration. Hearing isn’t as good as in vertebrates; it’s likely that the windy nature of the planet meant that sensitivity to sound wasn’t as useful, since the wind often drowns it out. In fact, the endostean ear is more effective at picking up vibrations in the ground, as in platysomes who often have their mouths pressed against the ground. Unlike airborne sound, sound is able to travel effectively through the earth even in windy conditions.
Inside the mouth are a number of taste receptors, including receptors on the roof of the mouth as well as the tip of the tongue. There are scent receptors in the respiratory slits in some groups, although smell is rarely a strong sense.
In addition to these senses, there are also pressure receptors in the skin throughout the body, as well as temperature and pain receptors. There are three balance organs in the head just behind and under the brain (one rear and two front), able to sense orientation, and most muscles have stretch receptors.
 |
| Evolution of Endostean eyes |
Reproduction
Endosteans have two sexes, although the males are much smaller and remain worm-like in form throughout their lives. The males mate by latching onto a female and slowly fusing to her, providing the female with a regular supply of sperm. Males tend to go inside the female's breathing slit; females probably originally had the sexual organ the males latch onto outside it, but it moved into the slit for protection.
In most species females "mate" by giving each other their male offspring. Some produce males asexually when their eggs haven't been fertilised, and females when their eggs have been fertalised; however, other Endostean groups will have to have mated before to birth any males.
Most Endosteans start out as worm-like larvae, metamorphosising as they mature, although some groups have fully-developed young. Larvae may or may not form a protective cocoon depending on the specific clade.
Young are grown in a uterus-like organ in the head, with a birth canal leading to the oesophagus. Eggs or live young go through the oesophagus and are expelled out of the mouth.







{kind=link}