Close to the eastern coast of the continent of Utiwa, near the equator, there is a region that consists largely of grassland during the windy season, and savannah and temporary forests during the still season. Located near a seasonal ocean, the region can get much wetter in comparison to the deserts further to the northwest, especially during the torrential rain in the transitions from the windy season to the still season. This allows much more life to thrive than in the more inland regions, although the winds don’t allow plants to grow too tall. The dominant plant life in this region consists mostly of yellow grass-like organisms and bamboo trees.
Floriavis maximus

Size: 15 cm long from tail to head, 15 – 20 cm wingspan
Diet: nectar, seeds
Habitat: savannas, trees
Reproduction: All unfertilised ova develop into males, fertilised ova develop into females. Give birth to undeveloped young, which are stored in breathing slit.
Due to the existence of flower-like structures on many
plants on Eurus, nectivores are fairly common on the planet, just as on Earth.
One such group of nectivores are the florivorids. They belong to a broader group
of flying animals called the saurorniths, which secondarily redeveloped flight
using the wings ancestral to endosteans, previously used for other functions
like temperature regulation, balance, and gliding.
To aid in their nectivorous lifestyle, florivorids have a
number of adaptations such as their long, thin beaks and the ability to hover.
Many saurorniths, florivorids included, have their wings fused together so all
four can act as a single pair. And while this usually comes at the cost of not
being able to hover as effectively, this isn’t the case for florivorids. To
combat this, they’re capable of beating their wings at an extremely rapid rate,
using elastic energy stored in their tendons and other connective tissues to
help sustaining this beating once it’s started.
Unlike most other florivorids, who inhabit the wind shadow forests where flowering vegetation is more common, Floriavis maximus prefers more open spaces. It is larger than most of its relatives, unable to rely on its small size to hide from predators as effectively, and has developed a venom it can inject from its claws for defence. With less need for camouflage than other florivorids they instead have bright blue scales for display, with most individuals preferring to select mates for male exchange that have a brighter colouration. Their scale colour is due to a pigment they can’t produce naturally which they can only obtain from certain plants, so it’s a good indicator of fitness.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Tetraptera
Clade: Viviparitores
Class: Saurornithes
Order: Plaudaciformes
Family: Florivoridae
Genus: Floriavis
Species: F. maximus
Palapus litus

Diet: Tiny basal insect-like holocelypheans
Habitat: Burrows in savannah and grasslands
Reproduction: Lay eggs. Offspring begin small and worm-like, living in their mother’s pouch after hatching. Females grow into their adult form in a cocoon.
With extremely strong winds half of the year, burrowing
animals are very common on Eurus. The family Xenoaspalacidae contains species
especially adapted to burrowing, allowing them to hide underground to shield
themselves from the harsh winds, as well as protecting them from predators. Palapus litus is a typical Xenoaspalacid
– or Eurus mole – with a short cylindrical body, short legs, and front limbs
well adapted for digging with large, curved spade-like claws.
The existence of Xenoaspalacids is an example of convergent
evolution. The Eurus mole has developed similar features to Earth moles due to
them both occupying a similar niche, and as a result being subjugated to
similar evolutionary pressures. This kind of convergent evolution is particularly
common on Eurus, or at least easier to see. The large animals of the planet
already have a similar body structure to Earth’s tetrapod vertebrates – they
are therefore affected by similar environmental pressures in similar ways.
Palapus litus spends more time out on the surface than other related species, and as such has retained the capacity for sight, lost in many other Eurus moles. Its legs are also relatively longer than some other species, allowing easier movement on the surface, and it has larger eyes. They usually only come out during the still season, however, spending most of the windy season in the shelter their burrows provide, hibernating during the worst parts of it.
TaxonomyTree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Clade: Ovata
Clade: Triodontes
Class: Marsupiata
Subclass: Pilata
Order: Brachypoda
Family: Xenoaspalacidae
Genus: Palapus
Species: P. litus
Platycastor orientalis

Size: 40 – 60 cm long (not including proboscis)
Diet: plants, Tmemichthid “fish”, tiny insect-like Holocelypheans
Habitat: coastal areas, retreat to pockets of saltwater on
the seafloor plains during the windy season
Reproduction: Lay eggs. Offspring begin small and worm-like, living in their mother’s pouch after hatching. Females grow into their adult form in a cocoon.
Platycastor belongs
to a group of animals called the Proboscates, characterised by the presence of
a long flexible tubular proboscis at the front of their body, used for feeding.
This proboscis is unsupported by bone, instead controlled and kept rigid by a
series of strong muscular hydrostats. This proboscis developed from the lower
tongue, rolled into a tube and fused at the top, but in many groups this is no
longer apparent. In a large number of Proboscates, like Platycastor, the proboscis is seamlessly attached to the skin on
the outside of the face and covered in hair, and can’t be retracted. More
primitive Proboscate groups, in contrast, keep the tubular tongue rolled up in
the mouth when not feeding.
Winds on Eurus can easily become strong enough to carry
small animals like Platycastor off
the ground. While many small animals hide from the wind by burrowing, others
solve this problem by developing a flat, squashed shape that allows the wind to
just pass over them. Platycastor,
with limbs ill suited for burrowing and a body poorly adapted to moving through
tunnels, is one such animal. As a result, they face less competition from
similarly sized animals during the windy season, many of which are dormant
during this period. Platycastor,
however, is able to remain active throughout the year.
They tend to live near bodies of water, with Platycastor orientalis preferring coastal areas. They can swim, but are more comfortable on land, using their long proboscis to catch sea “fish” in addition to gathering algae and seaweed-like organisms. Platycastor will also eat “insects”, both those on land and in the water. During the windy season, when the oceans shrink, they retreat to the small lakes left over, feeding on the sea animals trapped there. Fish are often forced to cluster closer together at this time of year, at least those who don’t migrate further offshore or hibernate in the soil, so they’re easier to come by.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Clade: Ovata
Clade: Triodontes
Class: Marsupiata
Subclass: Pilata
Order: Proboscata
Family: Platycastoridae
Genus: Platycastor
Species: P. orientalis
Conorhamphus
erythrus
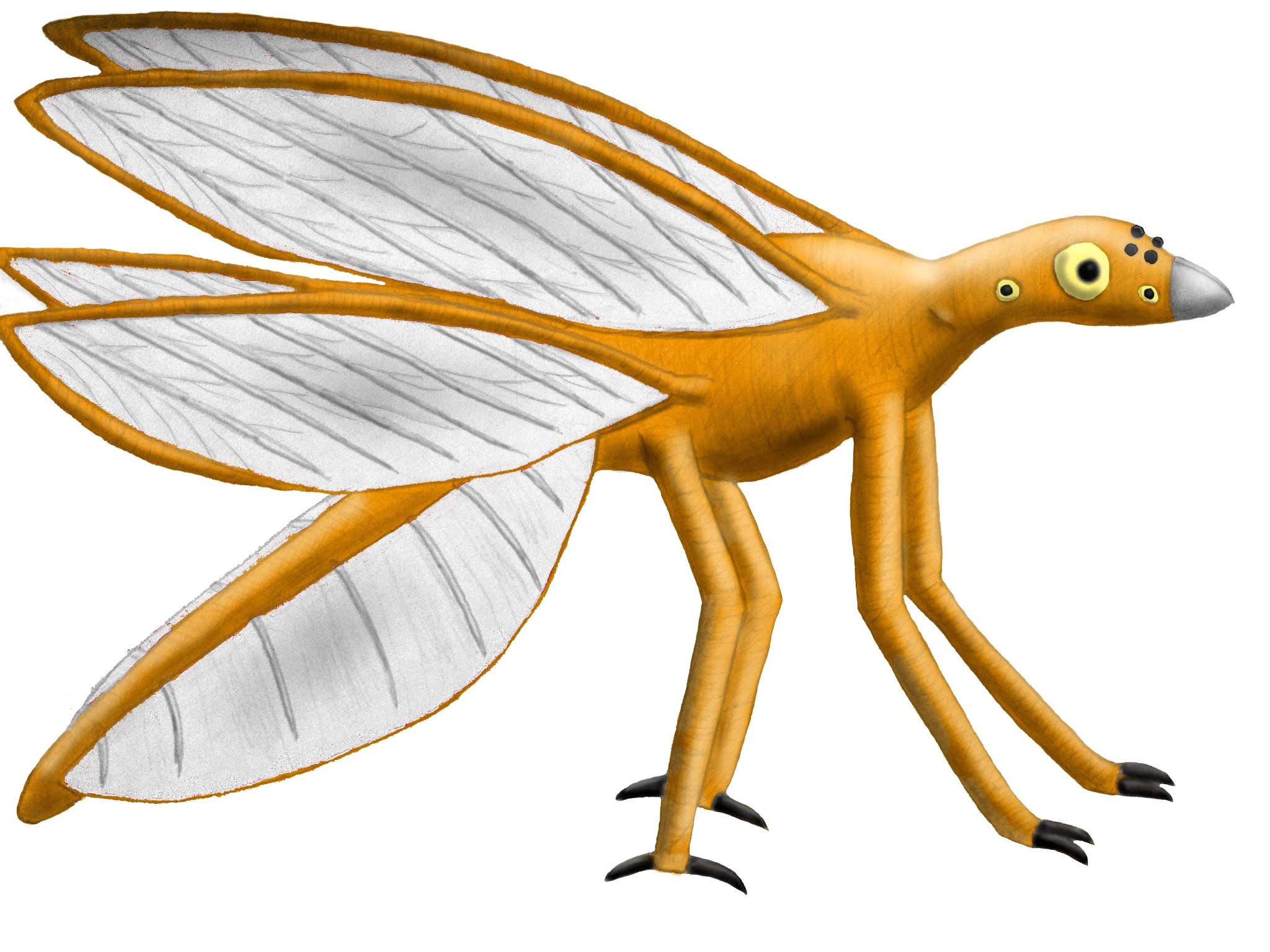
Size: 35 – 45 cm in length, including tail, 80 – 100 cm wingspan
Diet: seeds, fruit, insect-like Holocelypheans
Habitat: savannahs and grassland
Reproduction: All unfertilised ova develop into males, fertilised ova develop into females. Give birth to undeveloped young, which are stored in breathing slit.
Conorhamphus is a
much more basal Saurornith than Floriavis,
its four wings separate and with much longer legs. They belong to a group of
granivores and fruit eaters known as Dianisodactyls. While most members of this
group primarily inhabit rainshadow forests or the seasonally growing forests
that appear during the still season, Conorhamphus
prefers savannas, where it specialises in eating large seeds from bamboo trees.
It’s able to use its four wings to hover near the tops of the trees where these
seeds grow, staying relatively still as it feeds. In spite of the hovering
ability their two pairs of wings gives them, they don’t have the energy to
maintain hovering – or anything other than souring flight – for as long as Floriavis can.
Dianisodactyls are primarily characterised by the shape of
their rear feet, with their opposing toes allowing them to grasp tree stems.
Because of the shape of most bamboo trees, Dianisodactyls perch horizontally
rather than vertically. Their feet have a locking mechanism that allows them to
do this with relatively little energy, and they usually hold their bodies close
to the tree so there’s less torque, using their front limbs for stability.
Another feature shared by most Dianisodactyls is the
flexibility of their cartilaginous wing-supports; the series of “fingers” that
run along the edge of wing-supporting limb. These supports aren’t homologous to
digits, and have actually developed from hardened areas of the venation
ancestrally present in Tetrapteran wings. While in many Saurornith groups they
only provide support for the wing membrane, in the majority of Dianisodactyls they
have a flexible joint at the base. This allows them to fold up their wings when
they’re not in use. While most Saurorniths are able to hold their wings flat
against their body, the ability to make them smaller like this is rarer in
other taxa.
Conorhamphus can
unfurl its wings further than most Dianisodactyls, allowing it to modify their
aspect ratio. By unfurling its wings half way, they become much broader than
when they’re folded by have the same length; most Dianisodactyls only open
their wings slightly further than this. But past this point, the wings become
narrower and longer, until it reaches a point where they have the same width as
their folded state but are much longer. Conorhamphus
usually lacks the flexibility to extend its wings any further than this, but
this is much more than most of its relatives can extend their wings.
This ability to modify the aspect ratio of their wings
allows them to change it to suit the situation. The wings are extended to their
maximum extent during souring flight over longer distances, but to quickly
escape from predators low aspect ratio wings are favoured, so their wings
aren’t fully extended.
Other than this Cororhamphus
and other Dianisodactyls are fairly typical Saurorniths when compared to more
derived lineages like Florivorids. They have a beak, developed from the
mandibles of their ancestors, which are homologous to the inner mandibles of
Endosteans. This beak lacks the jointing of other Tetrapterans and has hardened
and become more beak-like. They lack the ability to chew food with this
structure, so food is ground up in a gizzard filled with chitinous plates. Unlike
birds, their beaks open to the side, as opposed to opening vertically.
Most of the digestive organs have moved back to the “tail”
of Saurorniths, actually an elongated and narrowed part of the torso, with the
area in front dedicated to flight muscles and the respiratory system. In
addition to their large, powerful flight muscles, which have a series of
anchors and pivots inside the rib cage for them to push and pull against, there
is a hydraulic pump providing these muscles with high pressure. Much like the
birds of Earth, the actual lung itself is quite small, but a series of air sacs
allows for the constant unidirectional flow of air both when breathing in and
when breathing out. Unlike birds, however, Saurornith air sacs can serve as
independently acting pumps.
Like many Tetrapterans, Saurorniths don’t lay eggs; they instead give birth to live young. Since this is quite energy intensive compared to egg laying for a flying species, and has the added disadvantage of weighing them down, pregnancy is short and the young they give birth to are very small and underdeveloped. Since many beak types don’t allow for the easy passage of larvae, in most species they burrow a hole through to the breathing slit and leave through there. The breathing slit also serves as a pouch for protecting their young as it grows, but they usually leave them behind in a nest or in their mate’s pouch if they need to leave for any extended period of time. Larvae are legless and beakless, and often blind.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Tetraptera
Clade: Viviparitores
Class: Saurornithes
Order: Dianisodactyliformes
Family: Conorhamphidae
Genus: Conorhamphus
Species: C. erythrus
Rhachioemys lambdanoton

Size: 15 – 35 cm
Diet: grass
Habitat: grasslands
Reproduction: egg-laying
This animal belongs to the order Acamptocormaria, which is
characterised by a particularly thick and sturdy shell-like skeleton. The
ribcage of Platysomes is already shell-like, lacking gaps between the rib
plates, but in most lineages these plates overlap to allow bending. However,
animals in this order have their ribs fused together so that the bones of their
torso act as one solid unit, providing strength at the cost of mobility.
As small, slow moving herbivores, this serves them well,
protecting them from predators. Their flattened body structure and stumpy legs
prevents them from running effectively, so having at least some form of
protection is essential to their survival. It is this adaptation that has
allowed the group to become so successful, with members of this diverse order
found throughout the planet.
Rhachioemys is a
non-Marsupiate, belonging to a distinct class called Septoculida, so named for
the presence of seven eyes; four major eyes and three smaller ocelli. While
these ocelli have been lost in many groups, Marsupiates included, they are
basal to Endosteans as a whole. Much more significant (but less immediately
visible) distinctions between the two classes are that Septoculids are cold
blooded, as well as the presence of chitinous serrations on their tongues.
These serrations help in the processing of food, lacking the acid glands of Marsupiates,
giving them an edge over other non-Marsupiates. Like Marsupiates they lay eggs,
but most species do little to care for their young.
Most Septoculids have a covering of scales to protect them
from the elements, since weather can get very extreme during the windy season
and the rainy transition to the still season. As relatively inactive animals
they will rarely attempt to seek cover, instead opting to endure these
conditions. Their eggs, too, are protected from harsh conditions, with a much
harder shell than those of Marsupiates, whose eggs don’t need strong shells
well protected in their pouch.
Rhachioemys has a
large, extendable and dextrous pair of tongues for obtaining food, allowing it
to reach for vegetation without having to move its body. It will use its
serrated tongues to grind up food before it even enters the animal’s mouth, at
which point it will bring the loose pre-processed vegetation into its mouth for
further chewing or just swallow it.
The animal has numerous hair-like sensory organs near its
mouth, characteristic of the order Acamptocormaria, which have developed from
hydrostatic extensions of the muscular oral surface. These are able to move
independently and are used to search for suitable food. Not only are they
highly sensitive to touch, but these organs possess taste receptors. They are
also able to extend their two frontmost eyes out of their head, raising them up
on stalks, which is used to search for food. The two larger outer eyes are used
for spotting predators.
The sensory organs at the front of the mouth can be used to detect incoming winds, and before the winds get too heavy they will usually crouch down so they’re closer to the ground, often tucking their limbs under their body. Although, with their stubby legs, this doesn’t do too much to change their overall height, it gives them a more aerodynamic shape allowing the wind to pass straight over them as if they were just a slight bump in the ground.
TaxonomyTree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Clade: Ovata
Class: Septoculida
Order: Acamptocormaria
Family: Ptilemydidae
Genus: Rhachioemys
Magnoros magnoros

Size: 2.5 to 3.5 meter high
Diet: grass, sometimes wood
Habitat: grassland
Reproduction: egg-laying, store their larvae in a pouch, build cocoons during metamorphosis
These large grazing herbivores come from the scaly, hairless
group of Marsupiates characterised by their erect limb structure. Magnoridans,
specifically, can be distinguished from other Marsupiates by their elongated
heads, giving them a long proboscis supported by jointed skull bone. This
allows the mouths of larger Magnoridans to easily reach grass or water without
them having to bend their hefty bodies or, in some species, allows them to
reach high leaves or seeds. Those belonging to the family Magnoridae, like the
species Magnoros magnoros, have a
series of hard chitinous teeth inside these elongated mouths, allowing them to
use this large muscular organ to grind up hard fibrous plant matter.
Magnoros magnoros is
among the most widespread and populous Magnorids, and is physically larger than
species from related genera. Comparable to an elephant in size and weight, this
is the largest animal found along the eastern coast, as well as many of the
other areas its various different subspecies inhabit.
The mouth of Magnoros is very large and thick, giving plenty of room for the interior to be filled with thick crushing musculature for the chewing of food. A pattern of contractions and expansions of the muscles underneath the layer of skin covered in teeth takes place as the animal spends hours processing its food. Like most Marsupiates, their mouths have acidic glands, further aiding in mastication.
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Class: Marsupiata
Subclass: Lepidodermata
Order: Magnora
Family: Magnoridae
Genus: Magnoros
Species: M. magnoros
Chenolarus utivensis

Size: 30 – 45 cm from shoulder to the tip of the tail, 50 – 80 cm wingspan
Diet: Tmemichthid “fish”, aquatic plantlife
Habitat: coasts, usually near equatorial grassland
Reproduction: Viviparous. Give birth to undeveloped young, which are stored in the breathing slit.
These relatives of Florivorids use their hovering ability to eat Tmemichthid “fish” rather than nectar, hovering over the water and catching fish in their beaks using their long necks. Their limbs are longer and they spend a great deal more time on the ground than their relatives, their heavier bodies unable to support a lifestyle almost permanently in the air. On land they can assume both a bipedal and quadrupedal stance, though aren’t as agile as they are in the air. Their webbed limbs allow for a limited degree of swimming, and their light underside and darker back provides camouflage via countershading.
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Tetraptera
Clade: Viviparitores
Class: Saurornithes
Order: Plaudaciformes
Family: Allochenidae
Genus: Chenolarus
Hadroglossus glaucos

Size: 1.3 – 1.8 meters in height, while stood with limbs erect
Diet: leaves, seeds, grass
Habitat: savannas
Reproduction: egg-laying, store their larvae in a pouch, build cocoons during metamorphosis
For most animals, there is a strong selective pressure
towards a flat, curved body structure that allows wind to pass over the body.
This can be at odds with certain life strategies, like for example browsing
from trees. Euran bamboo trees, in particular, keep their seeds particularly
high up, which can be a source of nutrition for many animals. This can be
difficult to reach for animals that aren’t tall enough, but are too large to
climb the relatively fragile bamboo trees. So in such animals, height and flatness
are two competing evolutionary pressures.
One family, the cheiroglossids, have the best of both
worlds. They stand tall, with long legs they’re able to hold erect below them,
but the torso itself is relatively flat. This allows them to keep their body
very close to the ground if they have to, just by crouching down during times
of heavy wind. They can move around fairly easily even while they’re crouched
in this position.
Since most bamboo trees will only grow during the still
season, standing tall poses no problem during this time of year. During the
windy season, with fewer trees, they have no need to stand as tall, and are
relatively inactive during this time of year getting what little energy they
can from grass.
One other adaptation common to cheiroglossids is the
presence of an elongated tongue, used in feeding. The tips of these tongues
have a pad adapted for grasping food.
Hadroglossus is
stockier than other cheiroglossid genera, with a series of hard bony bumps on
their back serving to protect them from predators. Sexual selection likely also
played a role; although the males are small and worm-like, females do select
other females to mate with. Or more accurately, to exchange males with.
Hadroglossus lives
in large herds for protection against predators, and in-group competition is
common, with individuals engaging in head-butting contests. They are highly
intelligent animals, with singing ability affecting one’s position within the
group as much as fighting. Hadroglossus
uses the two breathing slits on either side of its head to produce these songs,
which usually have a very low pitched humming quality to them, although they
don’t engage in song as much as some other cheiroglossids.
Although Hadroglossus isn’t as swift a runner as other cheiroglossids, their size does offer a degree of protection.
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Class: Marsupiata
Subclass: Pilata
Order: Ungulapedes
Family: Cheiroglossidae
Genus: Hadroglossus
Dolichorhis cursor

Size: 1.2 – 1.5 meters
Diet: grass, leaves, softer wood
Habitat: savannas, grassland
Reproduction: egg-laying, store their larvae in a pouch, build cocoons during metamorphosis
A much smaller and leanly build relative of Magnoros, the species Dolichorhis cursor shares the extended face characteristic of the order Magnorida. Unlike Magnoros, it lacks the chitinous teeth
present along the oral lining of its relative, having instead developed teeth
along its two tongues.
Dolichoris is a
cursorial grazer, depending more on speed and the protection its herd offers
than larger magnoridans do. Although not able to change direction quickly, they
can gallop in a straight line at an impressively fast speed, and have a great
deal of endurance. To facilitate their running, they stand only at the tips of
their nails. These have developed into hooves, a feature that distinguishes
them from most other magnoridans, the larger members of which have their hefty
bodies supported by fleshy pads on the soles of their feet. These hooves have
developed independently from those of the distantly related ungulapeds.
The rear eyes are larger than the frontal eyes, used primarily to watch out for predators. Its position higher up on the body and away from the proboscis is ideal for this purpose. The smaller eye pair, meanwhile, is much closer to the mouth and is used to search for patches of vegetation on the ground nearby. They are mainly suited for short distance vision, and their ability to make out shapes at a distance of more than a couple meters is very poor. The rear pair, on the other hand, are much more adapted to long distance vision than close vision.
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Class: Marsupiata
Subclass: Lepidodermata
Order: Magnora
Family: Dolichorinidae
Genus: Dolichoris
Oxyodon aegialus

Size: 30 - 50 cm long
Diet: small animals
Habitat: coastal grasslands and savannah
Reproduction: egg-laying, store their larvae in a pouch, build cocoons during metamorphosis
Oxyodon is a
small, long bodied carnivore that lives in burrows to escape winds and hide
from larger predators. They tend to focus on small prey around the same size as
themselves or smaller, such as Xenoaspalacids as well as Tmemichthid “fish”.
They will also supplement their diet with whatever worm-like organisms they can
find in their burrows.
There are a number of adaptations Oxyodon and other members of the order Oxyodontia have related to
their carnivorous lifestyle. As well as a relatively short digestive tract
common in carnivores, their mouths are well adapted for catching prey and
piercing flesh, with a long, sharp beak and outer mandibles ending in very
sharp teeth. The beak is visible even when the animal has its mouth closed. The
beak and teeth are able to retain their sharpness by constant shedding, with a
new, sharp tooth growing underneath. For this reason they have a habit of
scratching their beaks against trees or rocks to maintain their sharpness.
A hole is also located at the front of the face to allow clear passage for sounds to reach their hearing organs. As nocturnal predators their hearing serves them well, and since true night only falls during the still season heavy winds don’t interfere with their hearing as much. The frontmost eyes are positioned close to the front of the head, giving them good binocular vision. With a second pair of eyes on the sides of their heads, they don’t sacrifice peripheral vision for this, allowing them to remain alert for predators.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Class: Marsupiata
Subclass: Pilata
Order: Oxyodontia
Family: Oxyodontidae
Genus: Oxyodon
Species: O. aegialus

Size: 1 meter tall
Diet: small to medium animals
Habitat: savannah, temporary bamboo tree forests
Reproduction: egg-laying, store their larvae in a pouch,
build cocoons during metamorphosis
These large solitary ambush predators can be found
throughout the more forested parts of the savannah, where trees provide them
with plenty of cover to hide from their prey. Unlike most scaled Marsupiates,
they’re more active during the still season when this cover exists, hibernating
in caves during the windy season.
With their thick, sturdy bodies and short legs, they’re
unable to affectively chase down prey, or even pounce for them from a concealed
position, so they instead catch prey with their quick, lashing tongue. The
upper tongue has a number of adaptations for this; in addition to its length
and greater extendibility, the tip is covered in a number of chitinous spines
capable of sheering flesh.
Because of their shells, the earlier researchers thought
they were related to Magnorans at first, but they seemed to have developed this
independently. Their shells and large builds protect them from predators like
Pterodromids and Monocerodon, who
they can’t effectively run from. All four eyes are positioned near the front of
their body for good depth perception, and with their protective shells they
don’t have as much need for peripheral vision to keep an eye out for predators.
They’re solitary and highly territorial, only coming
together to mate, and tend to have large ranges. In-species fighting is
accomplished with their tongues, and they often aim for the eyes to blind each
other or attempt to damage each other’s tongue. Fighting is done to compete for
territory, and an individual will usually give up before they get too injured.
The only times they exist in groups is after mating, where two females will occupy the same territory with the offspring they produced from each others’ male offspring. They work together to ensure the survival of their larvae, and protect them once the females enter their cocoons. They continue to look after their offspring for a short period after they emerge from their cocoons, until they’re able to fend for themselves. The mothers leave each other straight afterwards, but will keep the male offspring they produced with them in their pouch to trade the next time they mate.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Platysoma
Class: Marsupiata
Subclass: Lepidodermata
Order: Pyrocheles
Family: Onychoglossidae
Genus: Onychoglossus
Species: O. vitriolosus
Dihoplus savanna

Size: 1.5 meters high
Diet: meat, primarily large herbivores including ungulapedes
and magnorans
Habitat: savanna, grassland
Reproduction: viviparous, young resemble miniature adults
These large cursorial predators belong to a group of
carnivorous, bipedal Tetrapterans called Pterodromids. Their body plan is very
different from the flying Saurorniths they’re distantly related to, and they’re
much larger than them too. Unlike Saurorniths these animals are flightless, and
use their wings for stability instead.
As they run, they hold their wings at such an angle that the
air passing over them pushes them downwards, giving them more traction against
the ground. Not only does this allow them to run faster, but being pressed
securely on the ground this way keeps them from being blown away in the wind; a
common problem for a lot of animals during the heavier parts of the windy
season, even larger animals. While many adapt to this by having their bodies
pressed against the ground, this doesn’t work as well for fast moving predators
like Pterodromids.
While most Pterodromids are incapable of sustained flights,
their wings are powerful enough that they can sometimes use them to provide a
degree of forward thrust during running. This is used more often by smaller
species than larger ones, although the fairly large Dihoplus does do this to an extent.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Superclass: Tetraptera
Class: Pterodromida
Order: Unguloraptores
Family: Dihoplidae
Genus: Dihoplus
Species: D. savanna
Monocerodon velox

Size: over a meter at the shoulder
Diet: medium to large-sized herbivores
Habitat: savannah and grassland
Reproduction: egg-laying, store their larvae in a pouch, build cocoons during metamorphosis
A much larger Oxyodont than Oxyodon aegialus, Monocerodon
uses its conical, spear-like tooth to take down prey. Like other Oxyodonts it
has a hole at the front of its head to allow for better hearing, but it is a
pursuit predator that takes on much larger prey than most of its relatives, and
has long limbs better adapted for running. Unlike most members of its order,
whose limbs are spayed out to the side, Monocerodon
holds its limbs in an erect posture directly under its body.
Monocerodon isn’t
a pure pursuit predator and will also ambush prey if necessary, usually if the
prey is larger than they are. Some species depend more on ambush tactics than
others, with the tactics used depending largely on the environment and the
amount of cover. Those who live in savannas and grassland like Monocerodon velox chase down most of
their prey, whereas some wind shadow forest dwelling species are exclusively
ambush predators.
After getting close enough during a chase, they leap onto
their prey using their powerful hind legs, piercing down on them with their
large, sharp tooth. If this doesn’t kill them instantly, the animal will die
very shortly afterwards from the blood loss. Monocerodon is also able to use its front claws as a weapon, usually
when hunting smaller prey.
These animals live in small groups consisting of a few related individuals. The dominant member of the group is typically the oldest. Monoceradon is most active during the still season, although they still do hunt during the windy season, mostly focusing on smaller animals. Individuals will fatten themselves up towards the end of the still season so they can last the windy season without as much food, and they’ll spend much of their time during the windy season huddled together for protection.
Taxonomy
Tree: Eurovitae
Domain: Euronucleata
Kingdom: Oxytrophia
Phylum: Holocelyphea
Subphylum: Endostea
Class: Marsupiata
Subclass: Pilata
Order: Oxyodontia
Family: Alloischidae
Genus: Monocerodon
Species: M. velox












{kind=link}